吴思实验室eLife报道海马位置细胞形成theta sweeps和相位移动的神经机制
2023年5月17日,北京大学心理与认知科学学院、IDG麦戈文脑科学研究所、定量生物学中心、北大-清华生命科学联合中心吴思团队,在《eLife》期刊在线发表了题为“Firing rate adaptation affords place cell theta sweeps, phase precession and procession”的研究成果,利用连续吸引子神经网络(CANN)加上神经元的适应性反应(adaptation)对海马区位置细胞进行建模,阐明了神经生物学实验所发现的海马位置细胞形成theta sweeps和相位移动的机制。
在行为过程中实时的对过去进行回忆、并对现在和未来进行想象预测是生物智能的一项重要能力。一个在进化上非常保守,普遍存在于啮齿类动物,灵长类动物和人类大脑的皮层下脑区——海马区——被认为与这项认知功能相关。
实验上观测到,当老鼠运动时,海马区的神经元集体活动产生的场电位(Local field potential, LFP)会产生4-12hz周期性震荡信号,被称为theta波。早期研究发现,位置细胞只会在特定的theta波的相位发电。这个相位会随着动物穿过该细胞位置场的过程而逐步提前。这一现象被称为θ相位进动(图1A)。在神经元群体的水平上,单细胞的相位进动会令位置细胞群体按照动物通过其位置场的空间顺序依次发放,在一个theta周期内形成正向的活动序列。这样的正向theta序列(图1A)由于其开始于老鼠过去的位置而沿着老鼠的运动方向结束于未来可能到达的位置,所以被认为是起到展望未来的作用。这对一系列认知能力(如计划、想象力和决策)可能有用。
除了展望未来,智能的行为还需要实时回顾过去。比如当前获得了某种奖励,就需要将这种奖励与之前的行为进行联系,从而总结获得奖励的经验,形成更有意义的记忆。最近的实验研究发现,海马区位置细胞同样会表征theta反向序列,并且在每个theta周期中与theta正向序列共存,且交替出现。这种有趣的相位编码模式实现了交替出现的“回忆(反向)序列”和“前瞻(正向)序列”(图1B)。
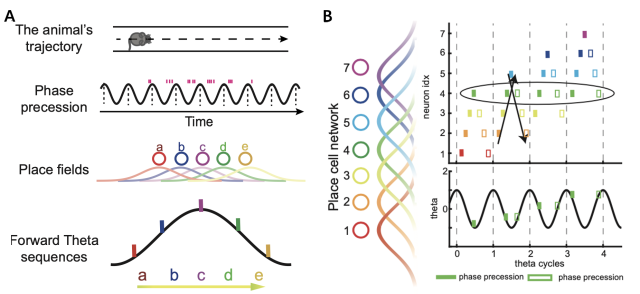
图1: theta相位进动以及theta序列示意图
虽然之前已经有大量的计算模型来研究产生相位进动和正向θ序列的神经机制,例如单细胞振荡模型和基于循环的活动扩散模型,但正向和逆向序列的交替共存,以及单神经元是如何实现相位进动和退动的共存的潜在神经机制仍不清楚。由于缺少可以研究的计算模型,该领域有很多问题仍待解决,比如:逆向序列是否与正向序列共享相同的潜在神经机制,还是它们是由不同的机制分别生成的?反向theta序列的神经活动强度是否与正向的θ序列相同?如果不是,在多大的程度上theta正向序列比theta反向序列的神经活动更强?又是什么样的机制导致了这两种序列的强度不一致?
为了回答这些问题,本文用包含自适应机制的连续吸引子神经网络来建模解释正向反向theta序列,以及单神经元层面的theta相位进动与退动的神经机制。通过研究网络的动力学,文章指出,自适应机制为网络产生的“波包”活动会引入的“内禀移动性”,即让网络不能维持稳态,而倾向于改变当前的状态。而当网络受到外界刺激时——比如当动物移动时,海马区位置细胞也许会接收到来自于内嗅皮层和其他感官皮层传来的“位置信号”——这种“内禀移动性”会与外界的刺激相互竞争,从而使网络的状态在不同参数下表现出不同的动力学行为。本文通过理论解析和数值模拟的方法表明,当自适应强度和外界输入的强度接近,网络的波包活动会一会被反馈抑制“推开”,一会被外界输入“拉回”,从而展现出在外界输入位置前后振荡的行为。
本文显示,模型处于震荡追踪状态时,其展现出的动力学行为正好可以解释海马区的相位编码以及theta序列:
1. 在群体层面,由于网络波包的震荡,编码动物位置附近对细胞会在一个theta周期内,先后以正向与反向顺序被序列式对激活,从而呈现出交替的“回忆(反向)序列”和“前瞻(正向)序列”(图2A)。
2. 在个体层面,当网络波包的中心通过神经元编码的位置时,该神经元会以最大概率发放动作电位。由于一个theta周期内,波包会一前一后两次扫过动物所处位置附近神经元编码的位置,所以每个神经元在一个theta周期内有两个发放时刻(相位),随着动物的移动,一个随时间不断前移(相位进动),一个随时间不断后移(相位后动)(图2B)。相位移动使得大脑把秒/分钟量级的行为经历压缩到了约200毫秒的theta周期,事件发生的具体时刻及事件之间的时间间隔被忽略掉了,系统只保留了事件发生的顺序,这体现了我们场景记忆的特点。
3. 通过调节不同的自适应强度m,该模型既可以复现出双峰细胞(既有相位进动也有相位退动)的编码模式也可以复现出单峰细胞(只有较强的相位进动)的活动模式。这一点的原理在于自适应机制影响了正向序列与反向序列的强度。反馈抑制是一个比神经激发慢得多的过程。这导致当波包正向振荡扫过某个神经元时,它产生的延迟反馈抑制将在波包反向振荡时才发起作用。这导致反向序列受到了来自正向序列的抑制。由于振荡追踪总是以正向序列开始,(因为初始运动是由外部输入触发的),因此即使反向序列也会用同样的机制抑制正向序列,抑制效果也是不对称的,正向序列总是强于反向序列。所以当m较小时,正向与反向的相互抑制作用弱,单神经元既能够做较强的相位进动也可以做较强的相位退动,可以解释实验上观察到的双峰细胞的相位编码模式。而当m较大时,反向序列受到严重的抑制,神经元的相位退动受到明显的抑制,最后展现出来只有较强的相位进动,而无明显的相位退动。这可以解释实验上观察到的单峰细胞的相位编码模式(图2C)。
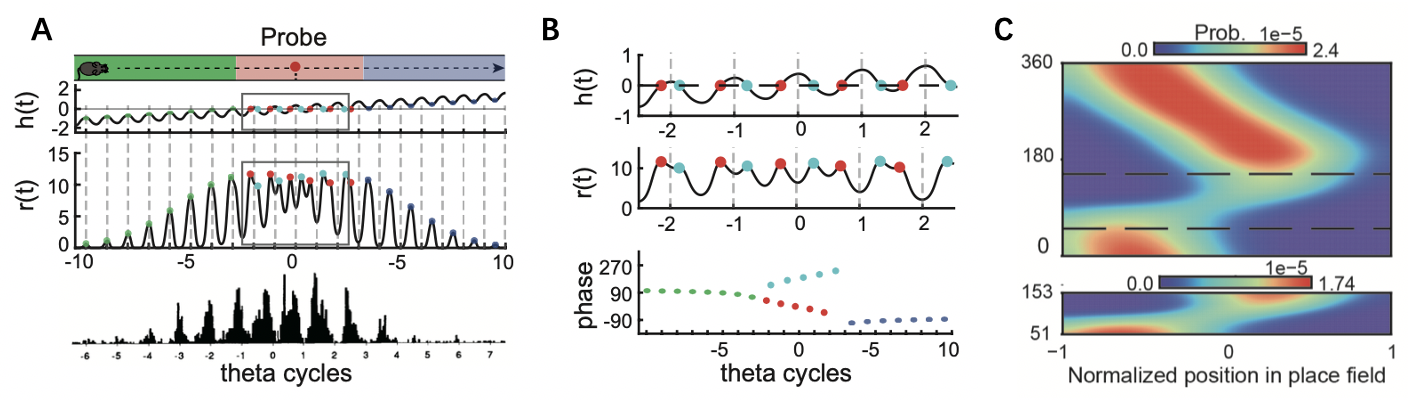
图2: 模型解释相位进动的原理
总结来说,本文通过建造神经网络模型,复现了海马区位置细胞的相位编码模式,以及产生了对应的“前瞻”和“回顾”序列。希望本研究能促进我们对海马时间编码的神经机制的理解,并为揭示海马时间编码的计算功能奠定基础。
原文链接
https://doi.org/10.7554/eLife.87055.1