Nikolai Axmacher:人类大脑中记忆印迹的再现、转化与损伤
撰文:宋永前,审核:王茜
2026年3月16日,受北京大学IDG麦戈文脑科学研究所邀请,来自德国波鸿鲁尔大学的Nikolai Axmacher教授在王克桢楼1113室作了题为“Reactivation, transformation, and impairment of memory traces in the human brain”的学术报告,王茜副研究员主持报告。

Axmacher 教授的研究致力于跨尺度揭示人类大脑中记忆印迹的形成与转化。其团队擅长将 7T fMRI、人类颅内脑电(iEEG)、人类单神经元记录等前沿技术,与人工智能的分析方法深度结合。在这场讲座中,Axmacher教授围绕“记忆印迹(Engram patterns)”这一核心概念,系统揭示了记忆印迹在人类大脑中从网络到单细胞层面的再现、转化机制,并深入探讨了心理创伤如何损伤这些珍贵的印迹。
引言:从“记忆过程”到“记忆内容”的范式转换
传统记忆研究往往侧重于大脑记住了“什么”(客观事件),但现代神经科学的核心挑战正转向探索大脑是“如何”记住这些事件的(独特记忆表征)。面对特定事件,大脑提取了多少感知觉细节,又提炼了多少高层语义?这种对事件独特的记忆表征,形成了独一无二且动态变化的记忆印迹 。
1. 记忆作为“再现”(Memory as Reactivation)
讲座的第一部分聚焦于记忆印迹的物理基础。动物模型表明,记忆印迹是特定事件及其情绪效价在海马或杏仁核细胞层面的神经表征;光遗传学重新激活这些细胞可诱发对应的记忆行为 。而在人类研究中,Axmacher 教授团队利用表征相似性分析(RSA),在宏观网络中成功提取了特异性的印迹模式。
在一项词汇-颜色联结记忆的 iEEG 研究中,只有当患者成功回忆起完整的“词汇-上下文”联结时,海马体才会精准重现编码时的特定时频神经模式。这证明海马体在记忆提取中扮演着重新绑定核心枢纽的角色(Staresina et al., 2016)。

图1. 海马体在词汇-上下文联结记忆提取中的时频印迹再现(Staresina et al., 2016)
那么,海马体如何与新皮层协作?团队利用虚拟现实(VR)结合 iEEG,观察患者在空间中寻找物体时的动态特征。数据揭示了记忆机制的时空动态变化:在回忆早期,海马体率先激活,重现“物体-空间”相绑定的关联索引;随后,外侧颞叶皮层(LTC)出现较晚的印迹再现,复原具体的“物体内容”。这表明,海马体首先提供索引,进而引导新皮层逐步重建感知觉内容(Pacheco Estefan et al., 2019)。
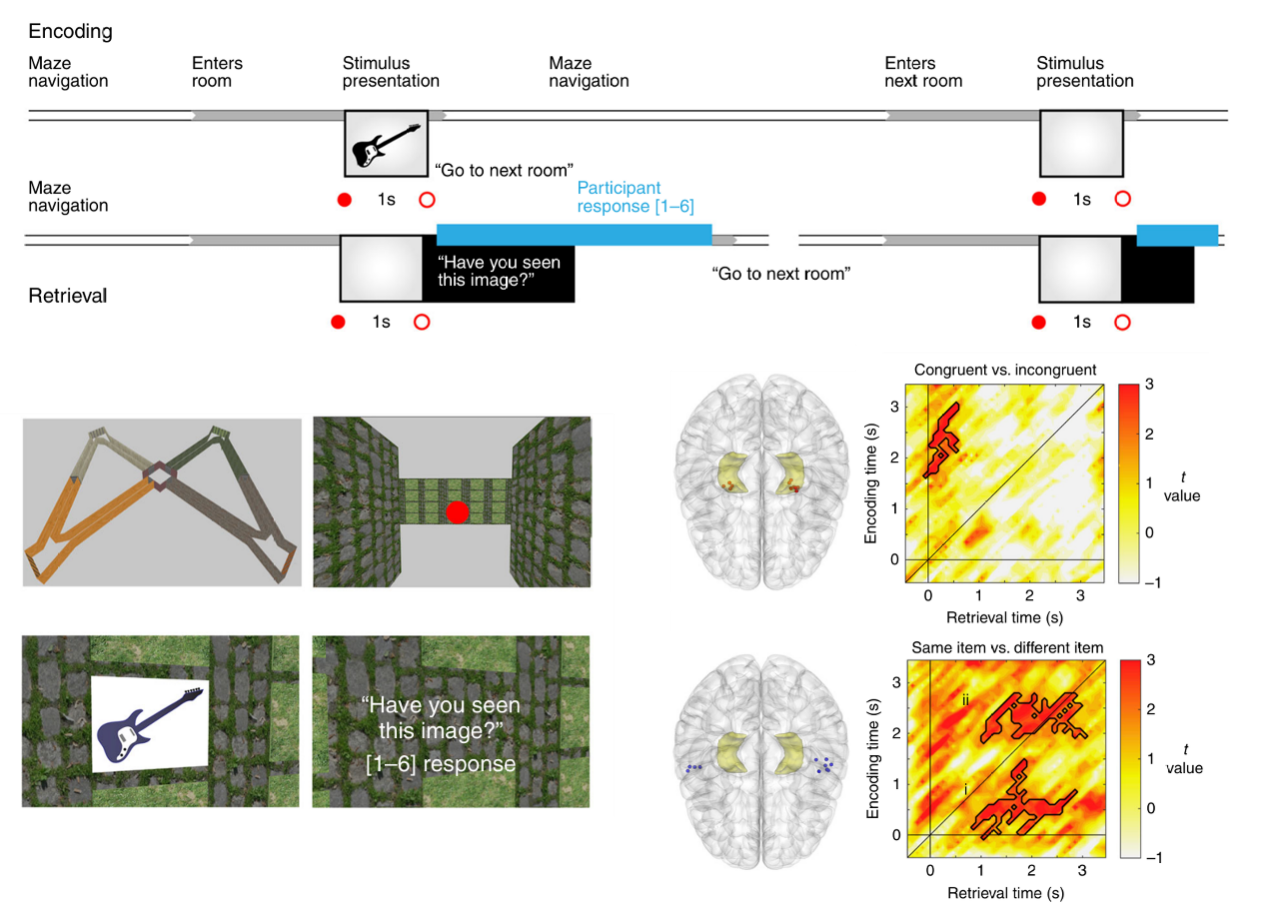
图2. 虚拟现实空间任务中海马体与新皮层的时空动态协作(Pacheco Estefan et al., 2019)
2. 记忆作为“转化”(Memory as Transformation)
传统观点常将工作记忆视为感知觉的静态延长,但 Axmacher 教授的研究指出,记忆实际上是一个高度动态的“转化”过程 。
研究团队引入深度神经网络(DNN)来量化短时记忆维持阶段的表征格式。结果发现,在记忆维持早期,大脑神经活动与 DNN 深层(如 fc6, fc7)高度吻合,主要提取高级视觉信息;而在维持后期,神经模式转为与纯粹的语义矩阵对齐 。这说明大脑对信息的加工进一步抽象化,并行维持了多种维度的表征格式,完成了从视觉到概念的实质性转化 (Liu et al., 2020)。
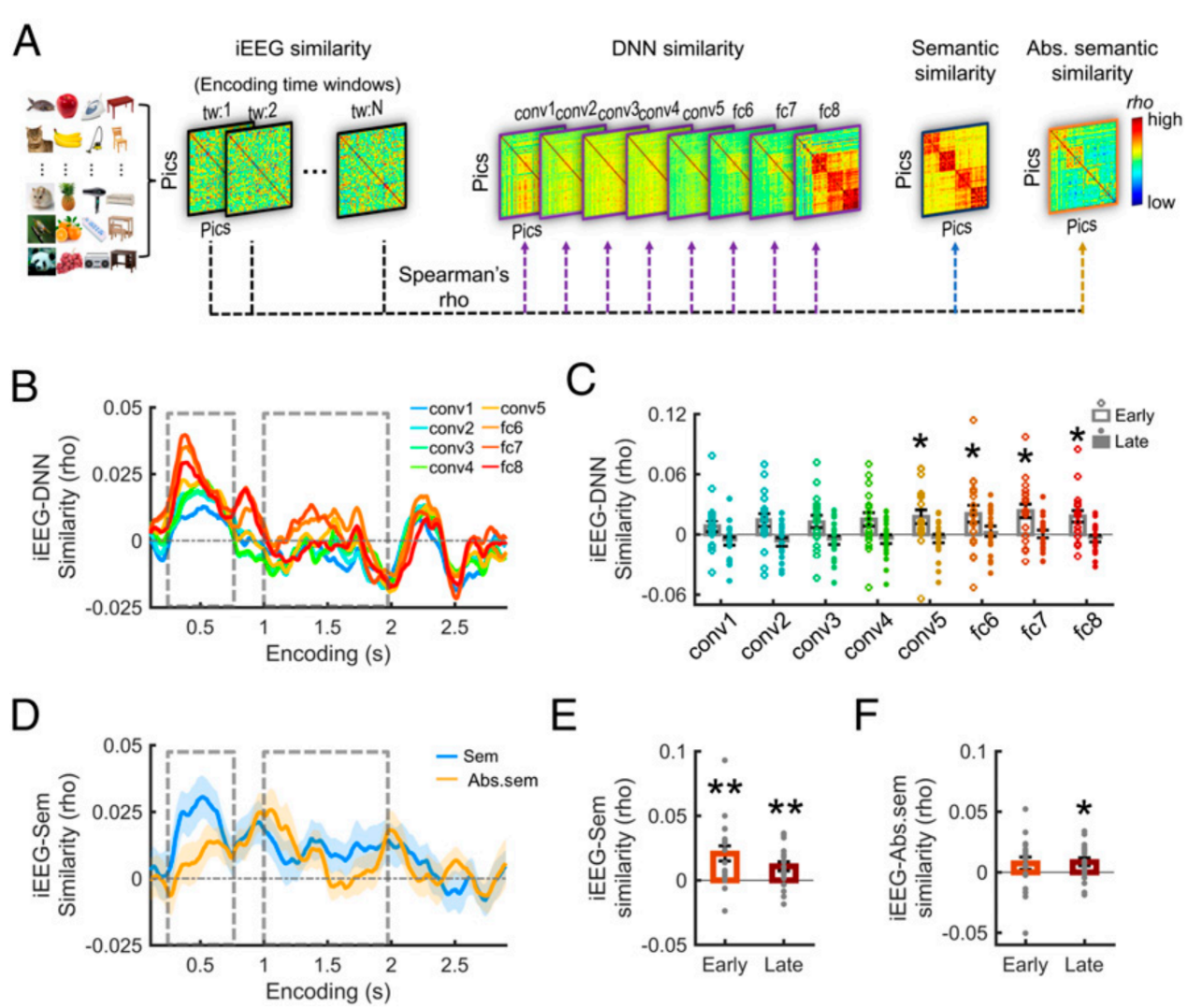
图3. 结合 DNN 揭示短时记忆维持期表征格式从高级视觉向语义信息的转化 (Liu et al., 2020)
此外,在多项目记忆任务中,只有包含反馈回路的循环神经网络(如 BL-Net 和 CORnet)才能成功拟合维持期的神经表征,前馈网络(如 AlexNet)则无法做到。这说明工作记忆中的信息不是简单的、静态的信息留存(前馈网络能做到的),而是一个在神经网络回路中不断循环、反馈和动态重塑的活动过程(Pacheco-Estefan et al., 2024)。
单细胞记录进一步证实了这一点:在工作记忆维持期,单纯负责编码“类别”或“位置”等独立特征的神经元减少,而同时加工这两者的“联合细胞(Conjunctive cells)”比例较编码阶段激增 76% 。工作记忆不仅是信息的“储存箱”,更是将其转化为复杂联合印迹的“加工厂” 。
3. 调和悖论:再现与转化的空间分工
海马体的“再现”与工作记忆的“转化”看似矛盾,大脑如何平衡这种关系?Axmacher 教授用一项关于再认记忆的研究地化解了这一悖论。在一项再认任务的研究中,对于同一张图片的编码与提取阶段,通过分析编码-提取相似度(ERS),可以衡量再现与转化的收益:如果一个图像被成功记住了,它的 ERS 大于那些被遗忘的图像,这就说明“高度相似”有利于记忆。反过来看,如果一个图像被成功记住了,它的 ERS 小于被遗忘的图像,这就说明“高度差异”反而有利于记忆(Rau et al., 2025)。
研究结果揭示了大脑精妙的空间分工。前颞叶皮层(aTC)作为高级语义枢纽,再现占据主导。神经模式越接近原始状态,记忆越牢固。外侧顶叶皮层(LPC)作为注意力与操作平台,转化是关键。信息被重塑得越偏离原始格式,记忆越容易被成功提取。
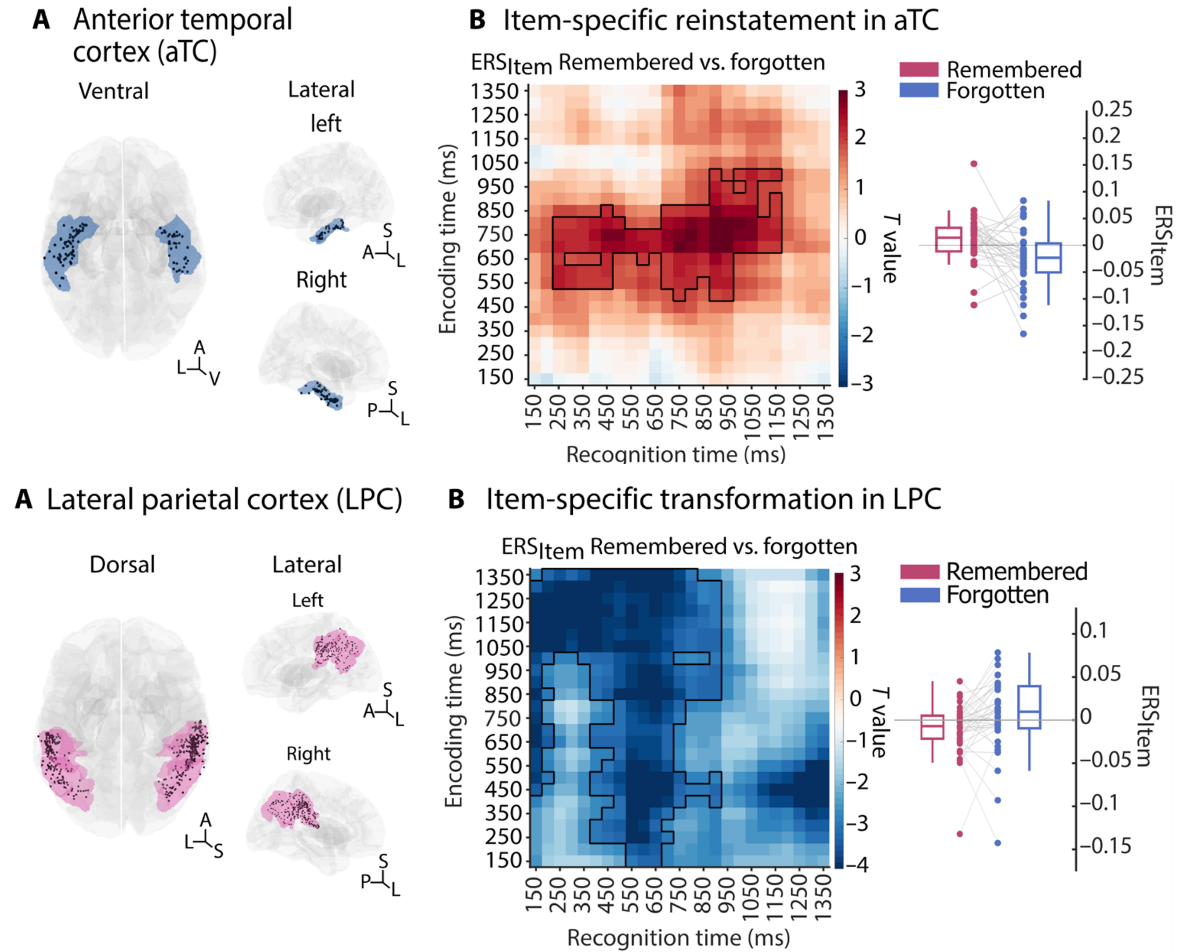
图4. 记忆再现与转化的空间分工(Rau et al., 2025)
4. 记忆的扭曲:压力、创伤与疾病预警
在讲座的后半部分,Axmacher 教授将上述计算表征工具应用于临床挑战。在极端压力或创伤下,记忆系统会发生严重扭曲,表现出生动但缺乏语义逻辑、极易被过度泛化(Overgeneralization)的特征 。
特里尔社会应激测试(TSST)结合 RSA 分析表明,社会心理压力不仅让人牢牢记住“核心物品”而忽略边缘细节,还会诱发杏仁核表征的过度泛化,使特定感知觉特征与恐惧情绪强行捆绑,导致表征空间的扭曲(Bierbrauer et al., 2021)。
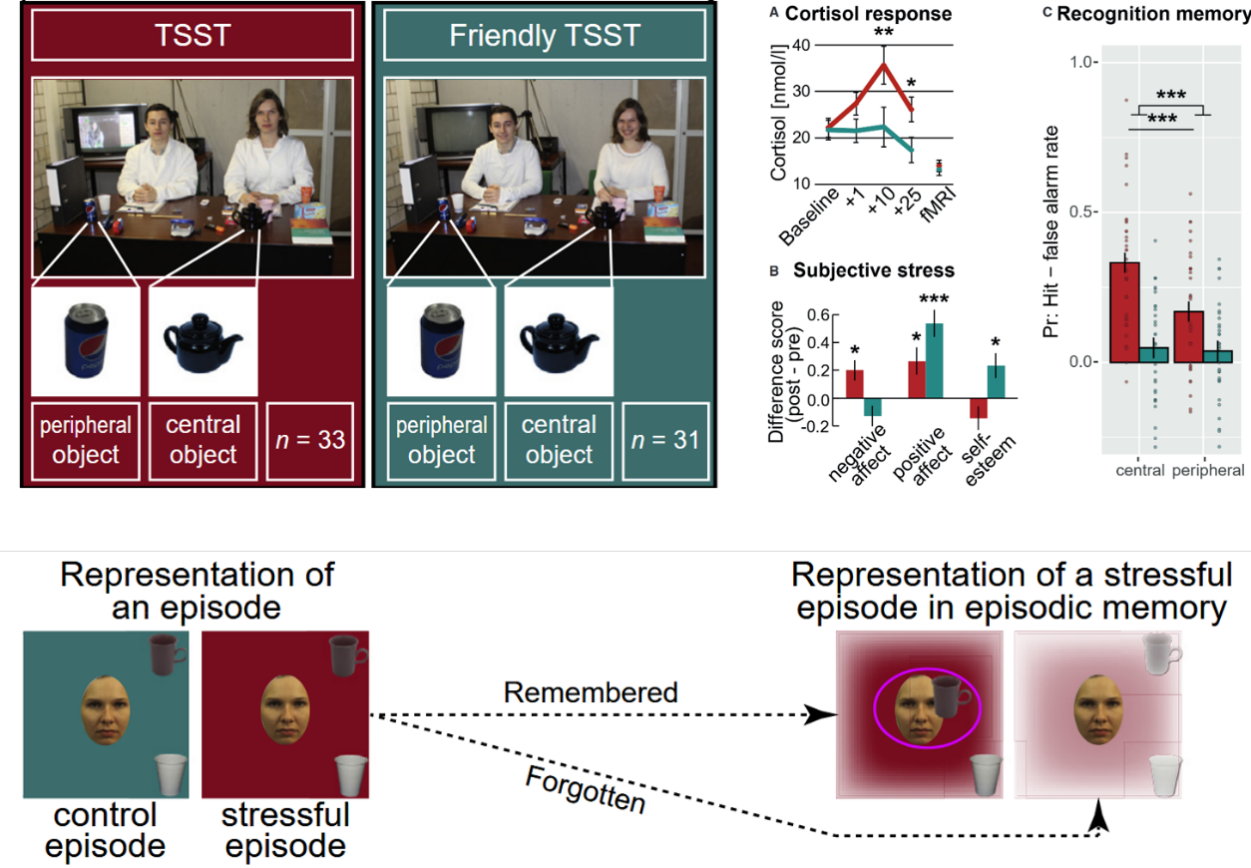
图5. 社会应激测试(TSST)揭示压力诱发感知特征与恐惧情绪绑定的过度泛化(Bierbrauer et al., 2021)
创伤电影范式进一步揭示了 PTSD “闪回”的神经机制:通过观看创伤视频,不同创伤事件在被试的神经表征上变得极其相似。这种过度泛化解释了为何 PTSD 患者极易被日常微小线索触发闪回,且真实发生闪回时的神经活动正是依赖于这些泛化的视觉表征 (Kobelt et al., 2024)。
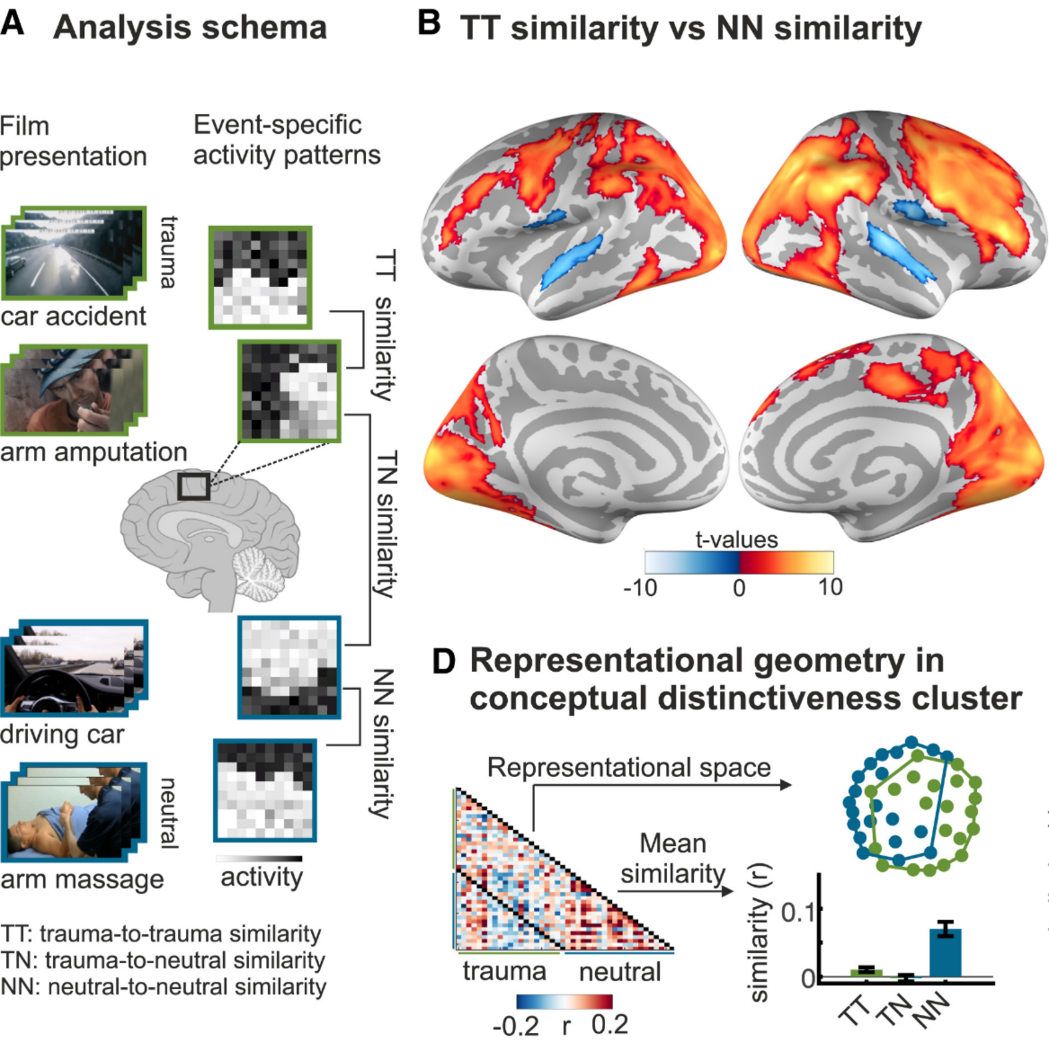
图6. 创伤电影范式下的神经模式相似度分析(Kobelt et al., 2024)
5. 空间导航与早期阿尔茨海默症(AD)
在 AD 症状出现前二十年,内嗅皮层的病理改变就已开始。研究发现,在平均年龄 22 岁、健康的 APOE-ε4 高风险基因携带者中,其内嗅皮层的“网格表征(Grid representations)”已出现显著衰退(Kunz et al., 2015)。
尽管他们在常规 VR 导航任务中表现正常,但这是因为大脑启动了海马体额外激活的代偿机制。通过剥夺视觉参照物的“纯路径整合(PPI)”极限测试,研究者成功消除了这种代偿伪装,暴露出高风险人群导航能力受限及策略改变的隐秘缺陷(Bierbrauer et al., 2020)。这一发现确立了受损的空间网格表征作为极早期 AD 预警标志物的巨大潜力。
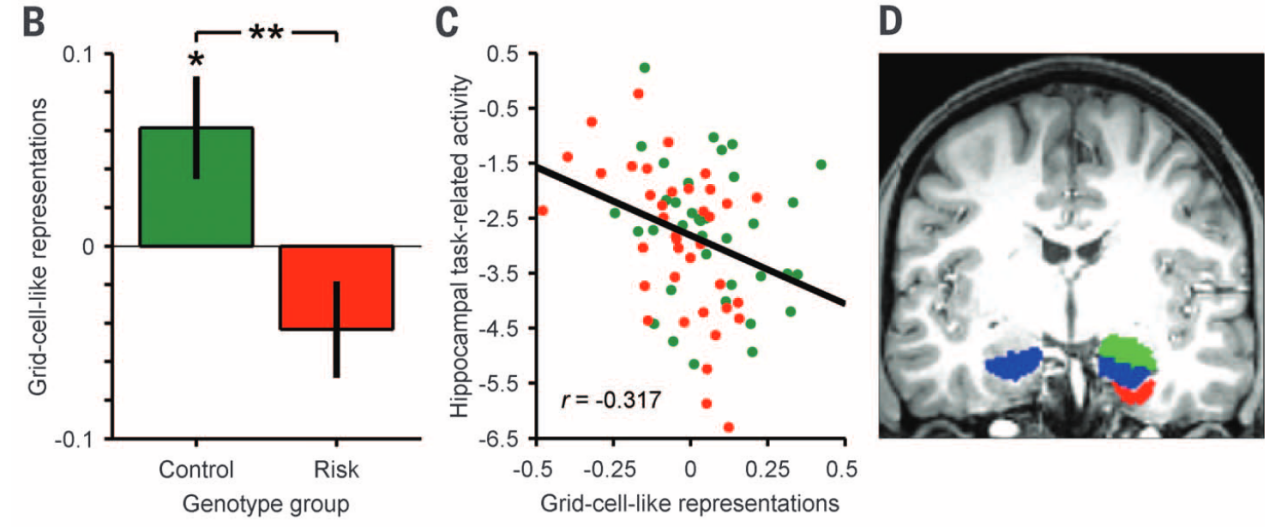
图7. 健康但高风险的 APOE-ε4 携带者内嗅皮层网格表征的极早期衰退(Kunz et al., 2015)
总结:跨越尺度的记忆解码
Axmacher 教授的讲座完成了一次从微观细胞到宏观行为、从基础理论到临床疾病的深度跨越 。其核心贡献可归纳为三点:
1. 印迹模式的跨尺度精确定位:借助多变量模式分析与 iEEG 等前沿手段,科学家现已能在网络和单细胞层面精准识别特定经验的记忆印迹 。
2. 再现与转化的二元统一:记忆的成功运作是一场精妙的交响乐,既需要“再现”原始痕迹(如海马体索引),又依赖于表征格式从视觉到高级语义的动态“转化”(如单细胞联合编码及顶叶皮层重塑) 。
3. 精准预警与临床洞察:解码这些印迹的底层逻辑,我们不仅能理解极端压力导致的感觉表征过度泛化(PTSD 闪回的神经根源),也能在 AD 悄然降临的二十年前,捕捉到内嗅皮层网格表征衰退的微弱求救信号 。
参考文献
1. Bierbrauer, A., Fellner, M.-C., Heinen, R., Wolf, O. T., & Axmacher, N. (2021). The memory trace of a stressful episode. Current Biology, 31(23), 5204-5213.e8.
2. Bierbrauer, A., Kunz, L., Gomes, C. A., Luhmann, M., Deuker, L., Getzmann, S., Wascher, E., Gajewski, P. D., Hengstler, J. G., Fernandez-Alvarez, M., Atienza, M., Cammisuli, D. M., Bonatti, F., Pruneti, C., Percesepe, A., Bellaali, Y., Hanseeuw, B., Strange, B. A., Cantero, J. L., & Axmacher, N. (2020). Unmasking selective path integration deficits in Alzheimer’s disease risk carriers. Science Advances, 6(35), eaba1394.
3. Kobelt, M., Waldhauser, G. T., Rupietta, A., Heinen, R., Rau, E. M. B., Kessler, H., & Axmacher, N. (2024). The memory trace of an intrusive trauma-analog episode. Current Biology, 34(8), 1657-1669.e5.
4. Kunz, L., Schröder, T. N., Lee, H., Montag, C., Lachmann, B., Sariyska, R., Reuter, M., Stirnberg, R., Stöcker, T., Messing-Floeter, P. C., Fell, J., Doeller, C. F., & Axmacher, N. (2015). Reduced grid-cell–like representations in adults at genetic risk for Alzheimer’s disease. Science, 350(6259), 430–433.
5. Liu, J., Zhang, H., Yu, T., Ni, D., Ren, L., Yang, Q., Lu, B., Wang, D., Heinen, R., Axmacher, N., & Xue, G. (2020). Stable maintenance of multiple representational formats in human visual short-term memory. Proceedings of the National Academy of Sciences, 117(51), 32329–32339.
6. Pacheco Estefan, D., Sánchez-Fibla, M., Duff, A., Principe, A., Rocamora, R., Zhang, H., Axmacher, N., & Verschure, P. F. M. J. (2019). Coordinated representational reinstatement in the human hippocampus and lateral temporal cortex during episodic memory retrieval. Nature Communications, 10(1), 2255.
7. Pacheco-Estefan, D., Fellner, M.-C., Kunz, L., Zhang, H., Reinacher, P., Roy, C., Brandt, A., Schulze-Bonhage, A., Yang, L., Wang, S., Liu, J., Xue, G., & Axmacher, N. (2024). Maintenance and transformation of representational formats during working memory prioritization. Nature Communications, 15(1), 8234.
8. Rau, E. M. B., Fellner, M.-C., Heinen, R., Zhang, H., Yin, Q., Vahidi, P., Kobelt, M., Asano, E., Kim-McManus, O., Sattar, S., Lin, J. J., Auguste, K. I., Chang, E. F., King-Stephens, D., Weber, P. B., Laxer, K. D., Knight, R. T., Johnson, E. L., Ofen, N., & Axmacher, N. (2025). Reinstatement and transformation of memory traces for recognition. Science Advances, 11(8), eadp9336.
9. Staresina, B. P., Michelmann, S., Bonnefond, M., Jensen, O., Axmacher, N., & Fell, J. (2016). Hippocampal pattern completion is linked to gamma power increases and alpha power decreases during recollection. eLife, 5, e17397.