Bryan Strange:不确定性与压缩——人类海马如何帮助自身形成记忆
撰稿:赵志睿 审核:王茜
2026年3月24日,受北京大学IDG麦戈文脑科学研究所邀请,来自西班牙马德里理工大学的Bryan Strange教授在王克桢楼1113室作了题为“Uncertainty and compression—how the human hippocampus helps itself make memories”的学术报告,王茜副研究员主持报告。

引言
在快速变化的世界中,海马(hippocampus)始终承担着记忆形成的关键任务。为了满足这一需求,它必须采取一种计算上高效的策略,在压缩感觉信息的同时适应环境中的不确定性。Strange教授的团队致力于使用脑磁图(MEG)、人类颅内记录和功能性磁共振成像(fMRI)等多模态方法研究人类记忆,尤其是情绪对记忆形成的影响。在本次讲座中,他围绕海马的两项核心功能——“准备(preparation)”和“压缩(compress)”,系统揭示了两者如何通过频率特异和状态依赖的神经动力学机制相耦合。
1 准备:海马动态通过涟漪活动适应刺激的可预测性
预测编码理论(predictive coding theory)认为,当背景(context)高度可预测或难以预测时,大脑会分别采取自上而下(top-down)或自下而上(bottom-up)的加工模式。视觉输入和预期的不匹配将导致预测误差(prediction error)的产生,并诱发相应的大脑活动。早在1966年,Vinogradova便在对清醒兔的电生理研究中发现,新异(novel)听觉刺激会引起海马CA1神经元的强烈发放,但随刺激的重复呈现,神经元的反应逐渐减弱,表现出典型的习惯化(habituation)现象(Vinogradova,2001)。而在之后的一项fMRI研究中,Strange团队也在人类左前部海马观察到了类似的习惯化效应(Strange et al.,1999)。
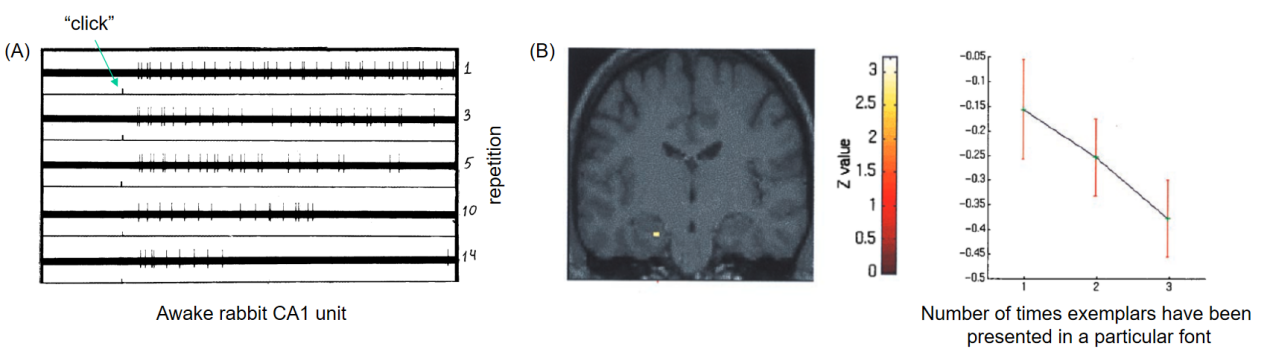
图1. 海马对新异刺激的反应及习惯化(Strange et al.,1999;Vinogradova,2001)
1.1 海马活动被环境的概率结构所调控
在上述研究中,刺激的新异性被定义为其相对新近性(即,该刺激最近是否出现过)。然而,海马是否同样会加工那些统计上异常的“oddball”刺激?采用经典的oddball范式,Dolan和Strange(2001)使用fMRI技术测量了被试海马和视觉皮层对不同类型oddball刺激的反应,这些刺激分别在知觉(perceptual oddball)、语义(semantic oddball)和情绪(emotional oddball)上偏离当前背景。结果显示,对三类oddball刺激,被试的左前部海马表现出一致的适应性激活,即在oddball刺激最初出现时作出响应,并随同类刺激的重复逐渐发生习惯化;与之相反,作为高级视觉皮层的双侧梭状回(fusiform gyrus)仅对知觉上的oddball刺激发生特异性反应,且未表现出跨实验阶段的适应。这意味着,海马具有一种学习异常情况发生规律的独特机制。
海马学习异常情况发生的可能性,这种机制实际上与Shannon的信息论(information theory)高度契合。在信息论框架中,惊异度(surprise)指某一事件发生的可能性,它被用来衡量事件发生时所携带的信息量,而熵(entropy)被用于量化某一事件的期望信息量,即事件发生之前的不确定性。因此,熵的概念似乎很好地捕捉了海马功能的本质。为探究海马是否对熵这一参数敏感,Strange团队与Friston教授合作,在一项fMRI研究中操纵了视觉刺激序列中不同刺激出现的相对概率比例(Strange et al.,2005)。与假设一致,被试前部海马对刺激序列的熵具有敏感性;反之,一个分布于双侧半球的皮层-丘脑网络的活动,则由每个具体刺激的惊异度所驱动。该结果表明,海马活动受到环境概率结构的调控,在事件发生前,海马便已经表征了该事件的预期信息量。Strange教授认为,海马对不可预测性的敏感性可能在知觉整合中发挥了关键作用,协助大脑在自下而上的感觉信息和自上而下的先验预期中建立平衡。
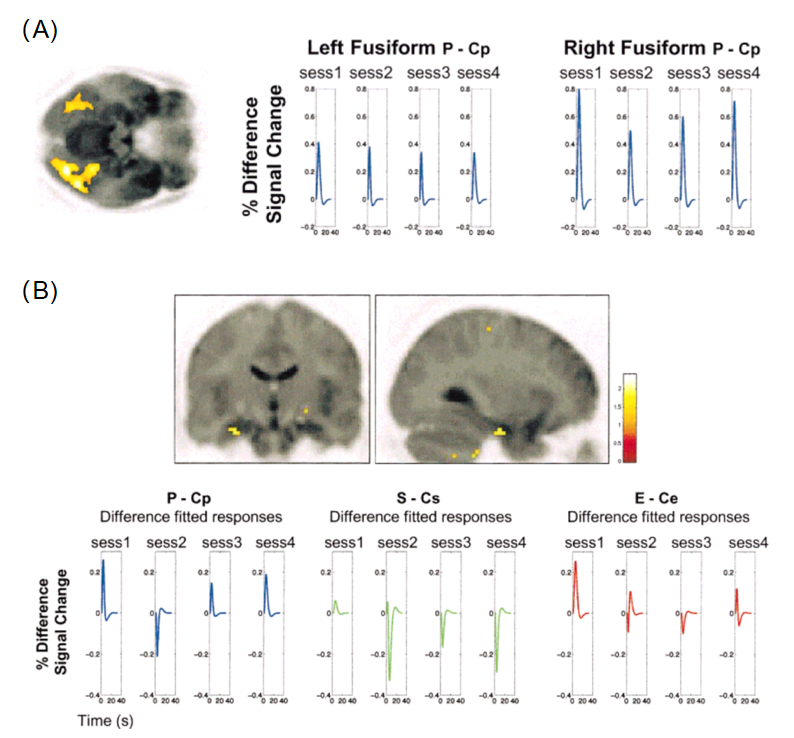
图2. 海马对三类oddball刺激表现出一致的习惯化效应(Dolan & Strange,2001)
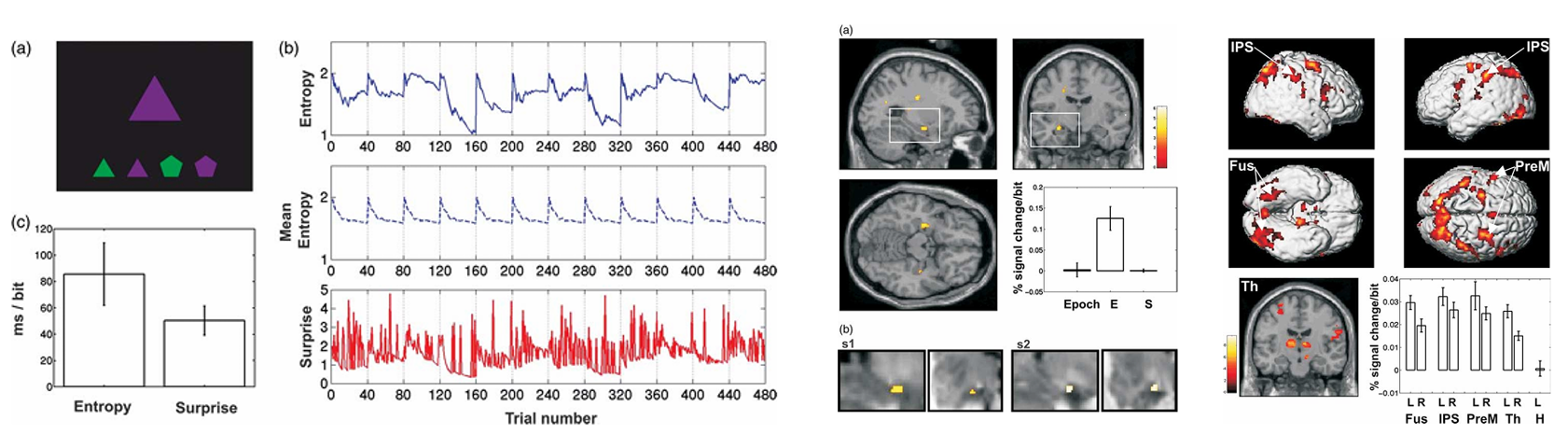
图3. 海马和皮层-丘脑网络分别对视觉刺激序列的熵和惊奇度敏感(Strange et al.,2005)
1.2 人类海马涟漪调节不确定性视觉背景中的皮层反应
然而,海马熵敏感性背后的神经机制是什么?理论上,倘若海马在预测未来事件中发挥了作用,其反应应发生在刺激出现之前,这是fMRI技术难以精确捕捉的。因此,在另一项研究中,Strange团队通过对人类海马和视觉皮层的颅内电生理记录,考察了一种特殊的神经活动:海马尖波涟漪(sharp-wave ripples,SWRs),它由尖波(大幅度的负极性活动,来源于CA1区锥体神经元顶端树突层的同步放电)和涟漪(140-200HZ的高频振荡,来源于CA3区兴奋性神经元与抑制性神经元之间的相互作用)共同组成。大量研究表明,SWRs不仅参与对过往经验的重放(replay),还能够表征环境中所有潜在可行但尚未实际经历的路径(Kay & Frank,2019)。Strange团队据此提出假设:在具有高度不确定性的高熵值情境中,个体需预测更多可能的未来状态,从而产生更多的SWRs。
结果显示,在高熵值情境下,刺激呈现前海马涟漪的发生率和持续时间都有所增加。进一步,刺激呈现前的海马涟漪会抑制枕叶皮层(occipital cortex)中与不确定性有关的γ频段活动,并调节刺激呈现后梭状回对惊异度的γ反应。与此同时,梭状回对高惊异度的γ反应也会通过该频段的定向连接反馈给海马(Frank et al., 2023)。这一回路表明,大脑中存在由海马涟漪介导的海马-皮层交互机制,以促进高精度预测误差的有效传播。
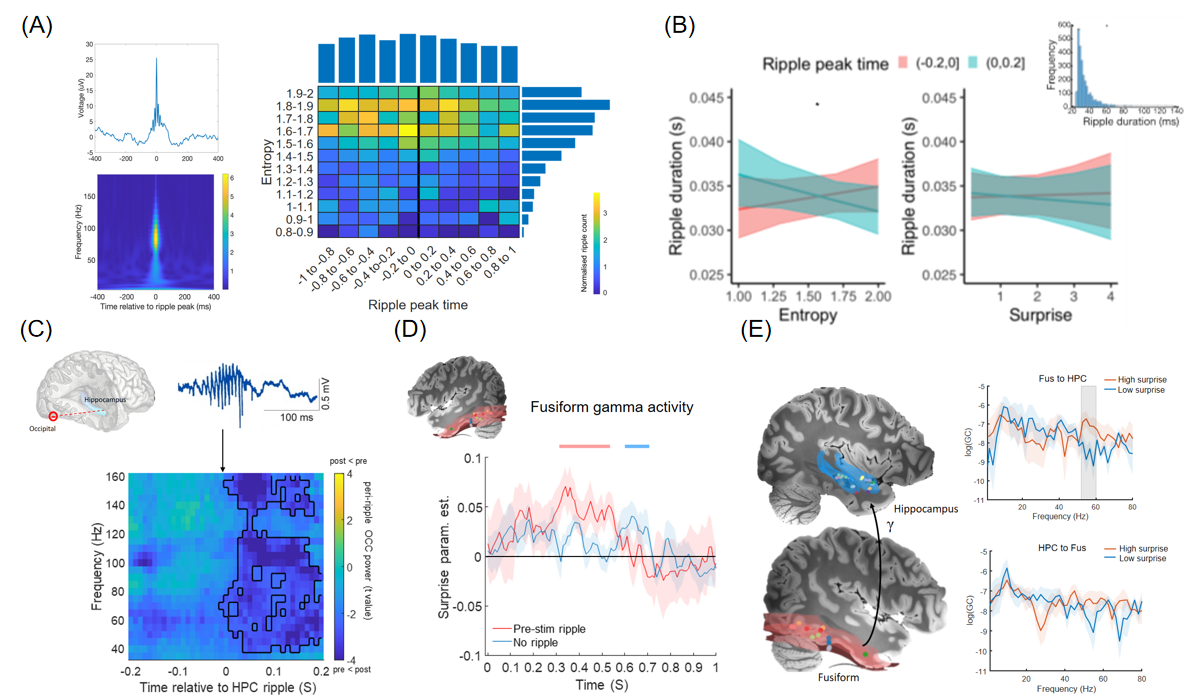
图4. 刺激呈现前的海马涟漪表征刺激的可预测性(Frank et al.,2023)
2 压缩:海马长轴根据空间频率压缩视觉信息
在讲座的后半部分,Strange教授介绍了海马长轴的功能组织。在啮齿类动物中,海马腹侧三分之一与背侧三分之二之间存在功能上的区分:沿海马长轴方向,其外在连接、受体表达及位置野大小均呈现出梯度变化。据此,Strange团队与Moser教授合作,于2014年提出了一个与海马长轴功能组织有关的新模型(Strange et al.,2014)。特别地,该模型认为啮齿类动物的腹侧海马可能在非条件化恐惧中发挥了选择性作用。
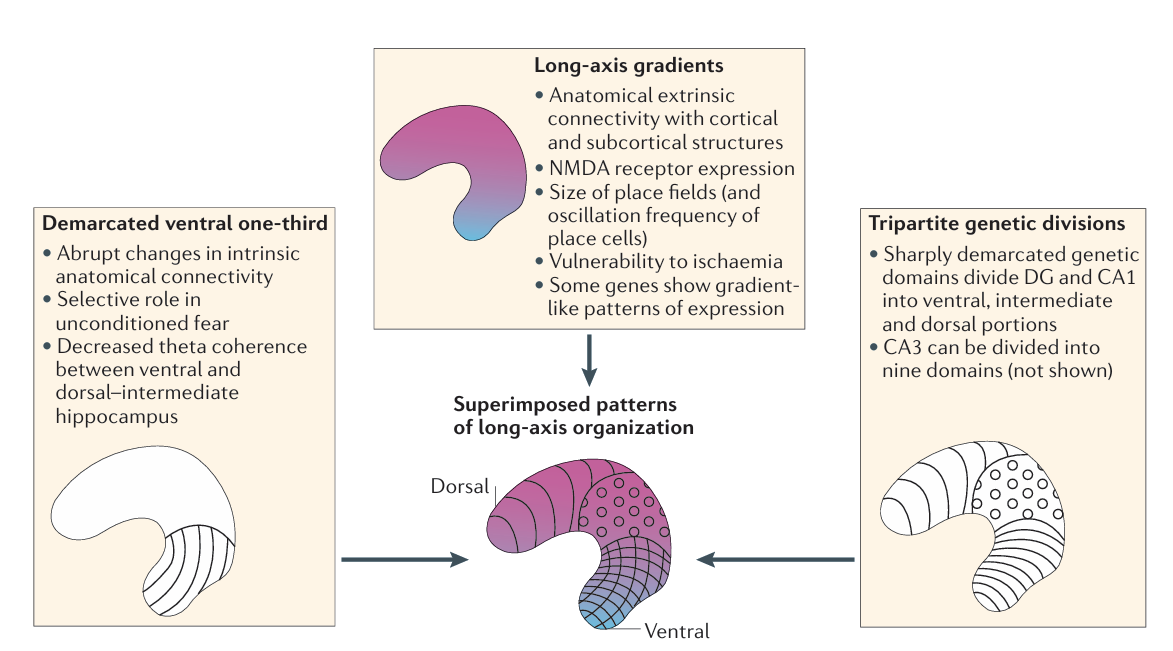
图5. 海马长轴的功能组织模型(Strange et al.,2014)
然而,在理解人类的记忆机制时,该模型存在一个根本性问题:作为灵长类动物,人类海马的功能组织可能显著区别于啮齿类动物。在一项关注杏仁核对情绪面孔反应的颅内脑电(intracranial electroencepholography,iEEG)研究中,Strange团队考察了人类海马对不同空间频率和不同情绪面孔的神经反应,试图检验人类前部海马(对应于啮齿类动物的腹侧海马)是否同样在非条件化恐惧中发挥作用(Méndez-Bértolo et al.,2016)。
出乎意料的是,Strange团队发现海马活动并不受面孔情绪的调节。与之相反,人类海马对面孔刺激的空间频率非常敏感,且其前部和后部分别对低空间频率和高空间频率的面孔刺激表现出选择性反应。Granger因果检验的结果显示,对全空间频率的正常面孔,存在一种从后部海马到前部海马的投射。这表明,人类海马长轴的不同部分分别对不同空间频率的面孔成分敏感。
进一步,颞中皮层向海马的投射是否也存在类似的空间频率敏感性?Strange团队在围嗅皮层(perirhinal cortex)和梭状回面孔区(fusiform face area)均记录到了对高空间频率敏感的γ活动。遗憾的是,他们未在海马旁皮层(parahippocampal cortex)记录到对低空间频率敏感的γ活动。未来的研究可以关注这种对空间频率的不同敏感性究竟源于何处、怎样产生,以及对面孔之外的其他刺激是否仍然成立。
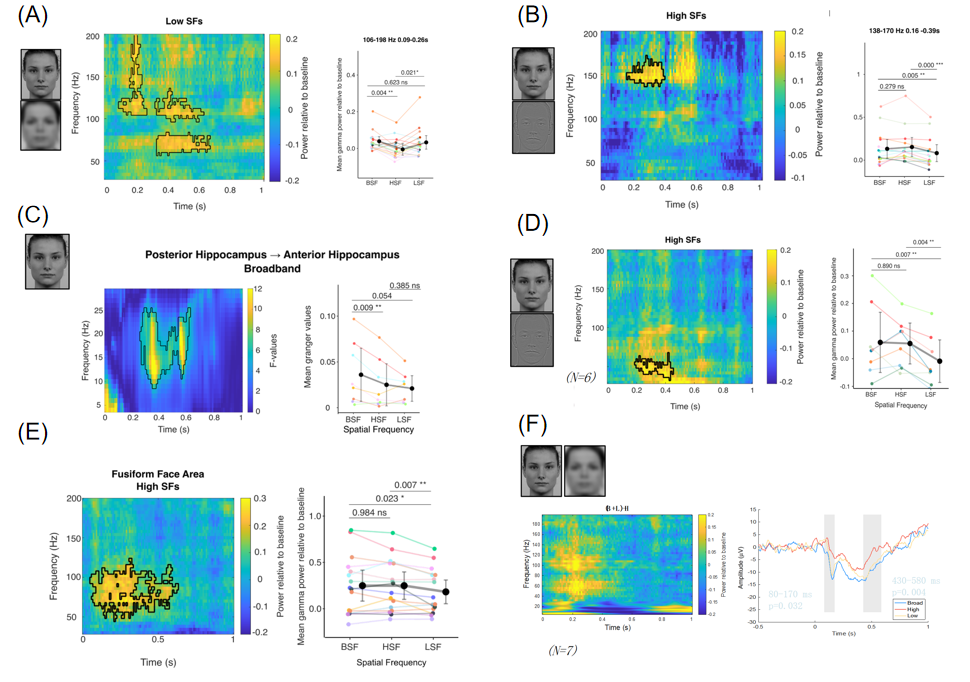
图6. 人类前部和后部海马分别对低和高空间频率的面孔刺激敏感(Méndez-Bértolo et al.,2016)
3 总结
最后,Stranger教授将本次讲座的内容类比为JPEG图片的压缩过程。JPEG图片的压缩先对图片进行颜色空间转换,然后进行离散余弦变换,将图片分解为不同空间频率成分,最后进行熵编码,对高频信息分配短码,低频信息分配长码。Stranger教授认为,人类海马可能首先对输入信息进行空间频率分解(前后部海马负责对不同空间频率成分的加工),然后通过类似熵编码的过程,根据信息的概率结构对刺激进行高效编码。总体而言,这些发现将海马的信息压缩、拓扑组织和预测信号整合到一个统一框架之中,共同支持人类的知觉和记忆形成。
参考文献
Frank, D., Moratti, S., Sarnthein, J., Li, N., Horn, A., Imbach, L., Stieglitz, L., Gil-Nagel, A., Toledano, R., Friston, K., & Strange, B.A. (2023). Human hippocampal ripples tune cortical responses in uncertain visual contexts. bioRxiv.
Kay, K., & Frank, L. M. (2019). Three brain states in the hippocampus and cortex. Hippocampus, 29(3), 184–238.
Méndez-Bértolo, C., Moratti, S., Toledano, R., Lopez-Sosa, F., Martínez-Alvarez, R., Mah, Y. H., Vuilleumier, P., Gil-Nagel, A., & Strange, B. A. (2016). A fast pathway for fear in human amygdala. Nature neuroscience, 19(8), 1041–1049.
Strange, B. A., & Dolan, R. J. (2001). Adaptive anterior hippocampal responses to oddball stimuli. Hippocampus, 11(6), 690–698.
Strange, B. A., Duggins, A., Penny, W., Dolan, R. J., & Friston, K. J. (2005). Information theory, novelty and hippocampal responses: Unpredicted or unpredictable? Neural Networks, 18(3), 225–230.
Strange, B. A., Fletcher, P. C., Henson, R. N., Friston, K. J., & Dolan, R. J. (1999). Segregating the functions of human hippocampus. PNAS, 96(7), 4034–4039.
Strange, B. A., Witter, M. P., Lein, E. S., & Moser, E. I. (2014). Functional organization of the hippocampal longitudinal axis. Nature reviews. Neuroscience, 15(10), 655–669.
Vinogradova, O. S. (2001). Hippocampus as comparator: Role of the two input and two output systems of the hippocampus in selection and registration of information. Hippocampus, 11(5), 578–598.