“人之初,首爱母”:饶毅实验室揭示从鼠到猴“亲母行为”的分子和细胞机理
【“人之初,性本善”是人类的良好愿望。
“人之初,首爱母”是正常哺乳动物的真实行为。

哺乳动物的第一个社会行为是亲母行为(maternal affiliation、maternal attachment)。通过研究多种动物(特别是包括猴在内)的亲母行为,揭示分子和细胞机理、特别是在多种动物中显示保守的机理,有助于理解人类的行为。
与其他社会行为、其他行为一样,亲母虽然是天经地义、却不是轻而易举,而是需要特定的分子、特定的神经细胞才能控制和实行。
出于好奇心,饶毅实验室长年对社会行为进行基础研究,希望在分子和细胞层面理解社会行为的机理,曾发现控制果蝇打架、老鼠求偶等社会行为的分子。
近日国际学术刊物发表他们延续十几年的一项研究,揭示控制“亲母行为”的分子和细胞机理。
他们发现,神经递质分子“五羟色胺”对亲母行为是必需的。如果动物婴儿脑内不能合成五羟色胺,亲母行为就会出现显著缺陷,幼崽不会很快去接近母亲,也不偏好母亲与其他雌性。而这些动物的嗅觉和视觉并无缺陷。
用遗传学和基因操纵证明,小鼠、大鼠、猴的亲母行为都需要脑内的五羟色胺。
这是人类第一次在同一篇论文用小鼠、大鼠到猴来自遗传学和分子生物学的证据证明一个基因与一种行为的关系。
他们进一步使用传统的电生理学、药理学和免疫组织化学,结合现代光学成像、化学遗传学、光遗传学、病毒遗传学等方法,发现五羟色胺来源于中缝核的神经元,它们投射到室旁核,释放的五羟色胺激活那里的催产素能神经元,通过社会性神经肽“催产素”,进而调控亲母行为的部分成分,在分子和细胞层面提供了机理解释。
这篇论文还提出一个可能普遍适用的机理:五羟色胺作为社会性神经肽的主控(master regulator)而调控哺乳动物多种社会行为】

众所周知,很多动物不是单独生活,而具有社会性。哺乳动物出生后的第一个社会行为是依附于母亲(所谓“亲母行为”,maternal affiliation、maternal attachment)。
亲母行为不仅对于婴儿获得营养、寻求保护等生存本能很重要,而且可能影响其成年后的行为。在人和灵长类,都有证据显示婴幼年缺乏亲母行为可以损害成年后的生活。即使母亲或看护者有虐待婴幼儿行为,婴幼儿也仍然表现亲近行为,更从反面说明亲近行为的重要性。
科学对于亲母行为的分子和细胞机理理解不够。神经肽催产素(OXT)和精氨酸-加压素(AVP)对社会行为很重要,被认为是“社会性神经肽”。它们参与小鼠的亲母行为,缺乏它们的时候,影响剥夺母亲引起的小鼠婴儿发送超声波(USVs)。但是,啮齿类亲母行为中OXT的具体功能有争议:有报道OXT基因敲除的小鼠幼崽减少发放USV,而另有报道外源OXT的加入可以减少大鼠幼崽发放USV。
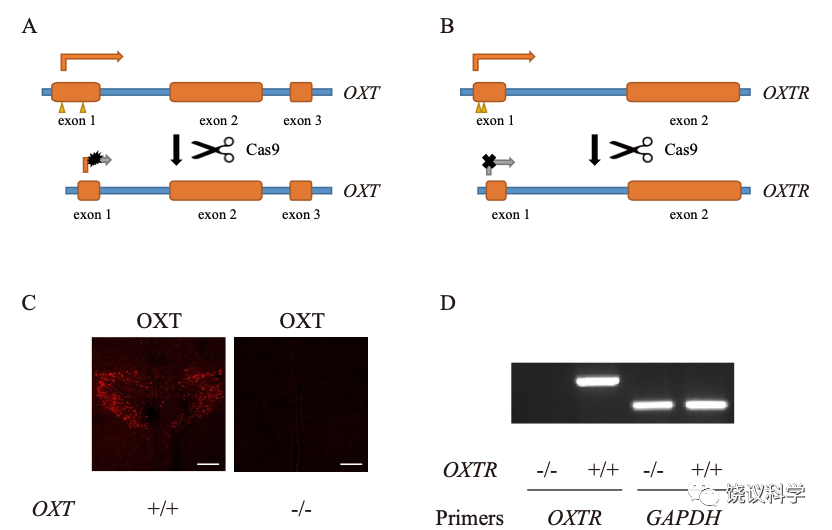
刘琰等制造了剔除“色氨酸水解酶2 ”(Tph2)基因的啮齿类和非人灵长类。这个酶为脑内合成五羟色胺(5-HT) 所必需。其突变后动物脑内不能合成五羟色胺。这种突变动物有助于研究和发现五羟色胺在亲和行为的作用。
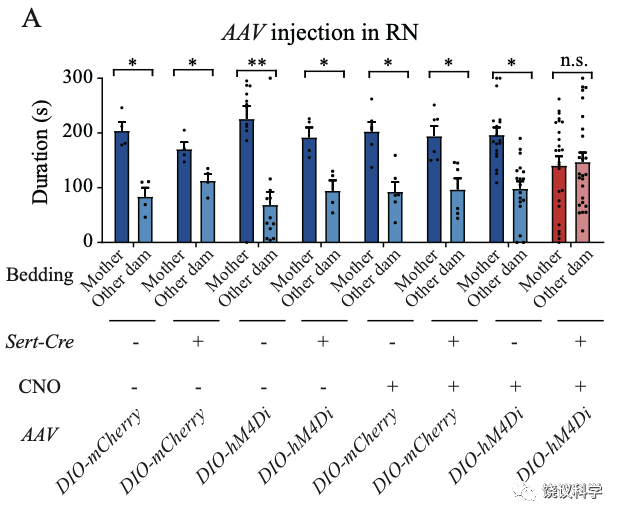
刘琰等进一步制造催产素OXT及其受体OXTR基因突变的大鼠,发现剔除它们的突变大鼠幼崽降低了对自己母亲气味相对其他母性气味的偏好。结合病毒遗传学、化学遗传学、药理学方法和钙成像以及行为分析,刘琰等发现给予OXT可以挽救缺失Tph2基因幼崽的母亲偏好。抑制中缝核(RN)的五羟色胺神经元导致幼崽缺乏母亲偏好,而这一缺陷可以通过激活室旁核(PVN)的催产素能神经元所挽救。 结果显示在亲和行为中,五羟色胺在催产素上游发挥功能。
缺乏五羟色胺(Tph2基因敲除)的小鼠幼崽缺乏亲母行为
2011年,刘琰等在《自然》发表的论文,报道五羟色胺参与成年雄性小鼠的性偏好。2013年,张莎莎和刘琰等在《美国科学院院刊》报道五羟色胺参与成年雌性小鼠的性偏好。
在研究过程中,他们意识到五羟色胺还参与多个其他的社会行为。从多个行为中,刘琰等挑选“亲母行为”进一步的详尽研究,因为它是哺乳动物出生后的第一个社会行为。
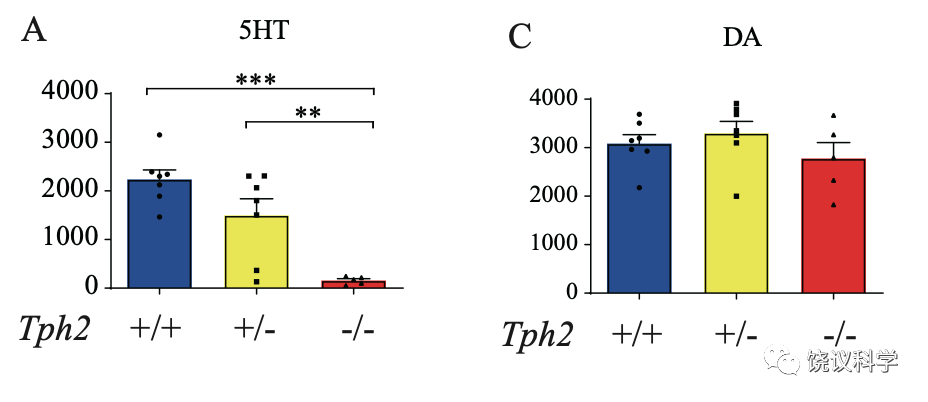
在亲母行为的研究中,刘琰等使用的小鼠幼崽的母亲都具有Tph2+/-基因型。而得到的幼崽有三种基因型(Tph2+/+,Tph2+/- or Tph2-/-),分别为野生型、杂合突变型、纯合突变型。通过高液相色谱检测它们的脑中的分子含量,可见分别含有正常量的五羟色胺、减量的五羟色胺、缺乏五羟色胺,而其他神经递质(例如多巴胺)含量在三种基因型的小鼠脑内没有显著差别。
刘琰等对幼崽进行超声检测后发现,在室温情况下,当幼崽与母亲分离后,会发出特定超声,这些超声被以前的科学家证明可以吸引母亲,求她们回来照顾幼崽。缺乏五羟色胺的Tph2-/-基因型小鼠幼崽发出的这类超声波少于其他两种基因型的小鼠幼崽。

因为母亲的体温高于室温,所以把母亲与幼崽分开后,幼崽可以感受温度的降低。为了排除温度的影响,刘琰等把小鼠放在类似母亲体温(35℃)的环境中,再分离母子,三种基因型的幼崽发放超声都降低,说明温度确实是一个因素。但仍然观察到母亲分离引起的超声发放在缺乏五羟色胺的幼崽少于其他幼崽,说明五羟色胺参与分离母亲引起的超声发放。
小鼠幼崽偏好自己母亲的气味。如果提供两种垫料,一种有母亲的气味、一种没有小鼠气味的干净垫料,正常幼崽偏好含母亲气味的垫料。如果一种垫料有母亲的气味,一种垫料有其他雌性的气味,正常幼崽偏好有自己母亲气味的垫料。
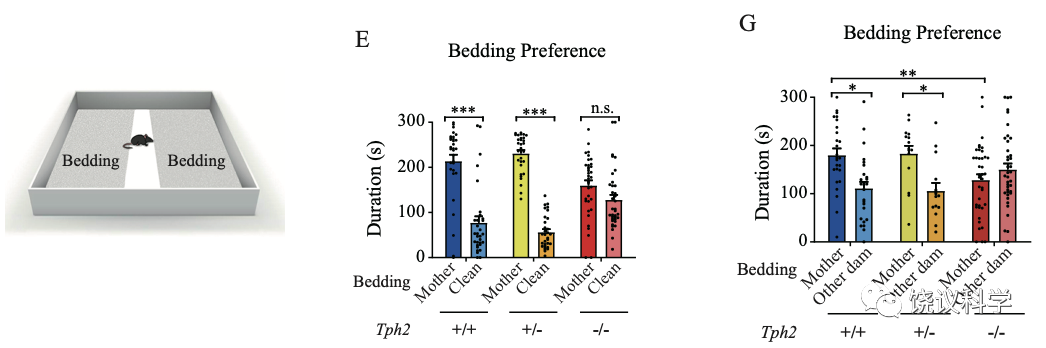
有五羟色胺的小鼠幼崽偏好母亲,而缺乏五羟色胺的小鼠幼崽对母亲的偏好显著降低。
这些行为偏好的缺陷,是否因为嗅觉缺陷所间接导致?刘琰等检测了小鼠幼崽对狐狸尿和薄荷的嗅觉。缺乏五羟色胺的小鼠幼崽与有五羟色胺的小鼠幼崽一样避开狐狸尿和薄荷气味。

这些结果说明脑内五羟色胺不参与一般嗅觉(薄荷)或社会嗅觉(狐狸尿)。
因此,实验结果支持脑内五羟色胺参与小鼠幼崽的亲母行为。
缺乏五羟色胺的大鼠幼崽有亲母行为缺陷
大鼠曾很长时间是神经生物学研究的常用动物。但因为1980年代发明的基因剔除技术很方便用于小鼠,而后来才能用于大鼠,使得神经生物学用小鼠更多、用大鼠更少。
CRISPR-CAS9基因修饰方法的诞生使得在包括大鼠在内的多种动植物的基因操纵都更容易。
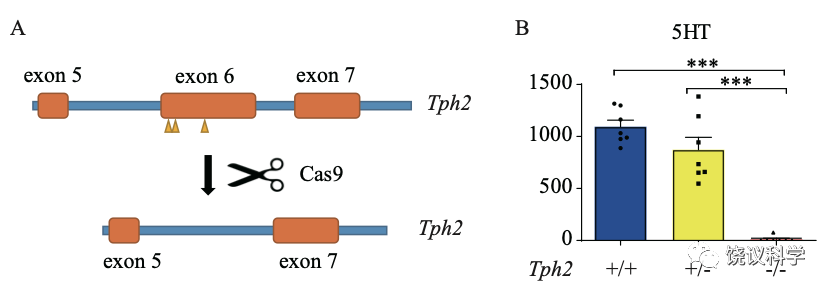
刘琰等用CRISPR/CAS9制造了剔除Tph2基因的大鼠突变种(Tph2-/-大鼠)。这种突变种的脑内五羟色胺也确实下降(而其他神经递质如多巴胺的水平不下降)。
与小鼠幼崽一样,大鼠幼崽被剥夺母亲后,也会发出超声。而与小鼠幼崽稍有不同的是,剥夺母亲后,再加回母亲后再次剥夺,大鼠幼崽第二次超声会更厉害,这种第二次剥夺导致的超声大于第一次剥夺的现象被称为“母亲增强作用”。这种现象在小鼠不是非常可靠,而在大鼠幼崽更可靠。这是用大鼠研究神经生物学的一个例子。
脑内含五羟色胺(Tph2+/+和基因型的)大鼠幼崽,不仅有第一次剥夺的超声发放,而且有母亲增强效应。而缺乏五羟色胺的大鼠幼崽,不仅第一次剥夺后超声发放减少,而且缺乏母亲增强效应。含五羟色胺,但含量低于正常(Tph2+/-基因型的)大鼠幼崽,表型居于正常和缺乏五羟色胺的大鼠幼崽之间,说明有剂量效应。

刘琰等验证缺乏五羟色胺的大鼠幼崽仍然能够发超声,因为温度提高到56℃ 或降低到4℃ 的时候,它们与含五羟色胺的大鼠幼崽一样发超声。
刘琰等用含气味的垫料做偏好实验时,含五羟色胺的大鼠幼崽在母亲和干净垫料之间偏好母亲、在自己的母亲和其他雌性之间偏好自己的母亲。而缺乏五羟色胺的基因突变大鼠幼崽失去这些对自己母亲的偏好。
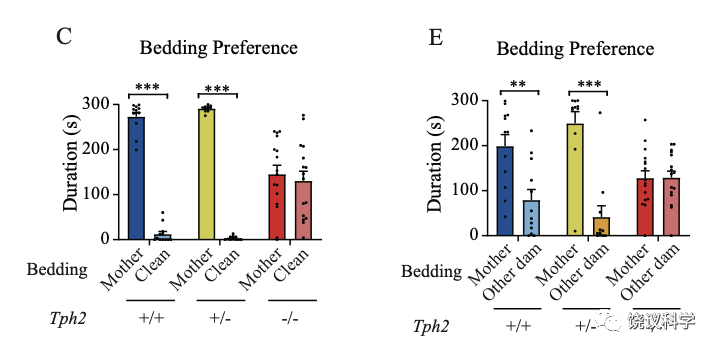
与小鼠幼崽一样,缺乏五羟色胺的大鼠幼崽没有嗅觉降低问题:同样避免狐狸尿和薄荷、喜欢牛奶。
以上实验说明小鼠和大鼠幼崽的亲母行为都需要有五羟色胺。
药理实验说明五羟色胺在出生后大鼠幼崽中发挥作用影响亲母行为
Tph2-/-的小鼠和大鼠幼崽都是在胚胎期开始就缺乏脑内五羟色胺。所以需要区分五羟色胺是在胚胎期起作用而后间接影响幼崽的行为,还是直接在幼崽脑内影响幼崽的亲母行为。刘琰等给正常幼崽注射药物p-chlorophenylalanine (pCPA) ,它可以抑制五羟色胺的合成,每千克体重注射200毫克的pCPA就可以在24小时内就可以几乎耗竭Tph2+/-大鼠幼崽脑内五羟色胺,而不影响其他神经递质如多巴胺的含量。
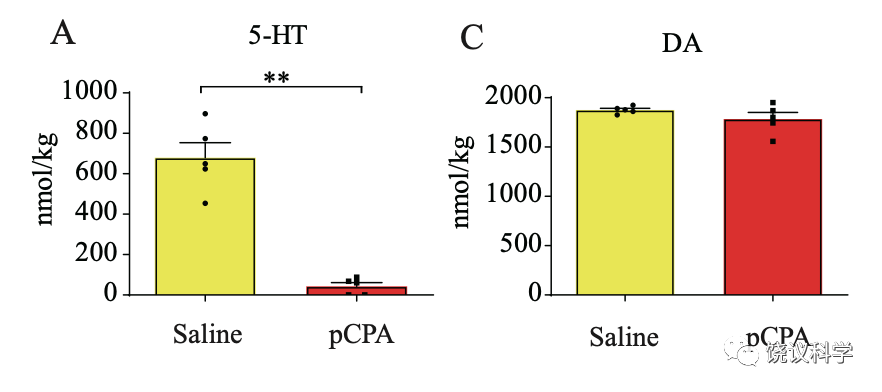
高压液相色谱分析显示,每千克体重注射200毫克的pCPA就可以在24小时内就可以让Tph2+/+大鼠幼崽脑内五羟色胺降低到其正常含量的31%,每千克体重注射600毫克的pCPA可以让Tph2+/+大鼠幼崽脑内五羟色胺降低到其正常含量的5%。
幼崽超声发送与脑内五羟色胺含量呈正相关。幼崽偏好母亲垫料也与脑内五羟色胺含量呈正相关。

为了进一步检测是否幼崽脑内的五羟色胺与行为有关,刘琰等给大鼠幼崽注射5-HTP,这是五羟色胺合成的前体分子,注射它可以绕过Tph2合成酶而提高Tph2-/-基因突变鼠脑内五羟色胺含量。在注射5-HTP后,Tph2-/-基因突变大鼠幼崽的亲母行为确实得到挽救。
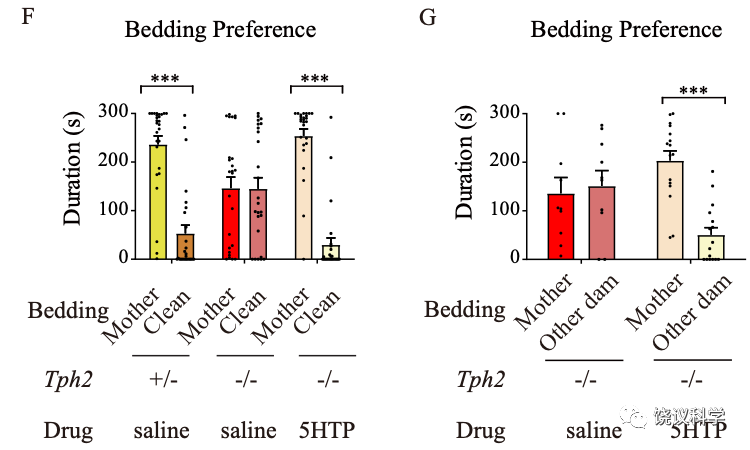
pCPA和5-HTP的两类药理学实验都支持五羟色胺在幼崽脑内参与亲母行为,而不是因为胚胎中的作用间接影响亲母行为。
缺乏五羟色胺的基因突变婴猴有亲母行为的缺陷
一个分子只影响啮齿类行为,不能知道它是否影响灵长类。
刘琰等与北京大学工学院的席建忠、昆明理工大学的陈永昌、牛昱宇等合作,用TALEN方法敲除了恒河猴的Tph2基因。
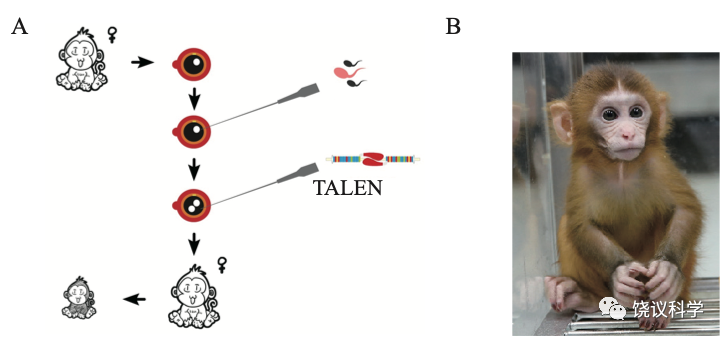
用高压液相色谱检测猴脑脊液的五羟色胺代谢产物(HIAA) ,确定它的含量降低,而多巴胺的含量没有降低。
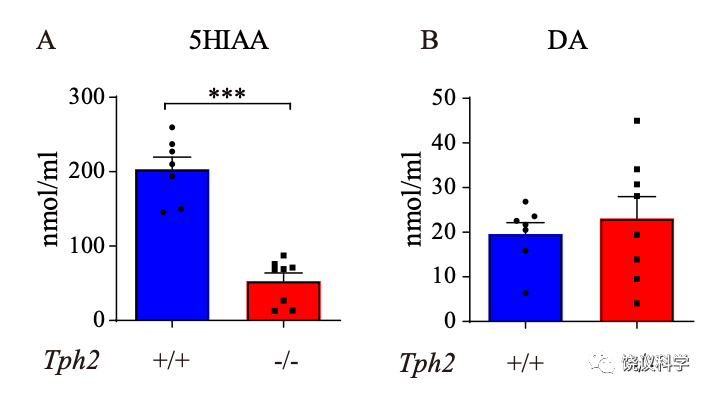
从80个移植中,得到9只Tph2基因敲除的(Tph2-/-)猴。它们由自己的母亲带大,其体重、运动与Tph2基因未敲除的猴无显著差异。
录像记录是普遍应用的分析婴猴亲母行为的方法。为了避免母亲行为干扰影响婴猴行为,刘琰等设计了去除母亲影响的两个实验。
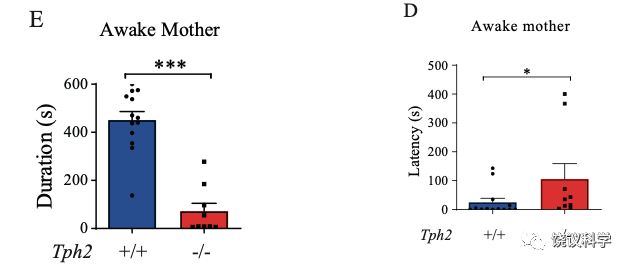
第一个实验室是短暂麻醉母亲后,把婴猴放进麻醉母亲的笼中,观察婴猴与母亲接触的时间。含五羟色胺的婴猴与麻醉母亲的接触时间远长于缺乏五羟色胺的婴猴。从婴猴进笼到接触母亲的潜伏期也是正常婴猴短于缺乏五羟色胺的婴猴。

第二个实验是让母亲和婴猴放在相邻的两个笼子里面,用栅栏相隔。正常婴猴走向母亲一侧的潜伏期短于缺乏五羟色胺的婴猴。正常婴猴在母亲一侧的时间长于缺乏五羟色胺的婴猴。
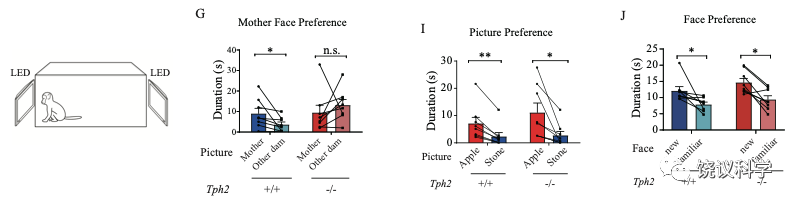
刘琰等还做了其他实验。例如,如果给婴猴看两张图片,一张是其母亲、一张是其他母亲,正常婴猴偏好自己母亲,而缺乏五羟色胺的婴猴不显示对自己母亲的偏好。但是如果给苹果和石头的图片,缺乏五羟色胺的婴猴与正常婴猴一样都偏好苹果。如果给一张新脸孔与一张旧脸孔图片,缺乏五羟色胺的婴猴与正常婴猴一样都偏好新脸孔。所以,缺乏五羟色胺可以影响对母亲的偏好,但同时没有迹象表明视觉受了影响、也不是记忆力的问题。
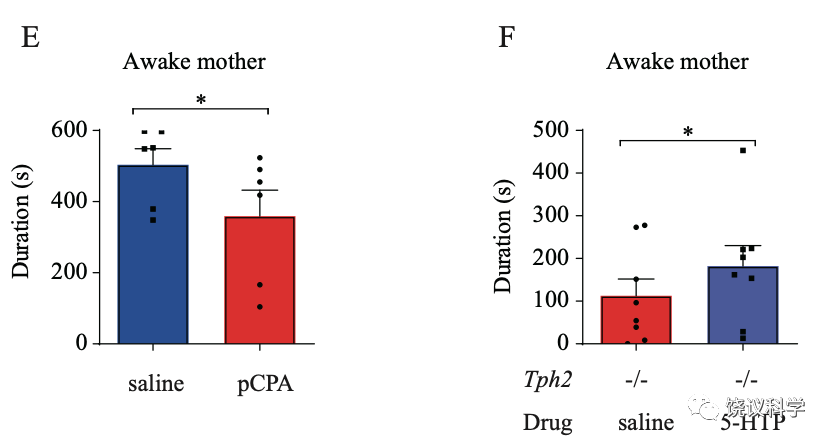
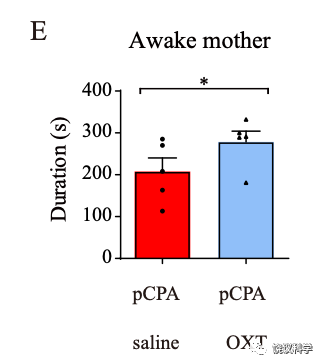
进一步的药理实验,用pCPA可以在正常猴导致五羟色胺降低,也出现亲母行为减少。而给Tph2-/-婴猴注射5-HTP,可以挽救其亲母行为的缺陷。这些实验结果支持五羟色胺是在婴猴脑中参与亲母行为,而不支持五羟色胺是在胚胎期间影响猴而间接影响出生后婴猴的亲母行为。
这些结果表明,从小鼠、大鼠到猴,五羟色胺都参与亲母行为。
母亲气味激活五羟色胺能神经元和催产素能神经元
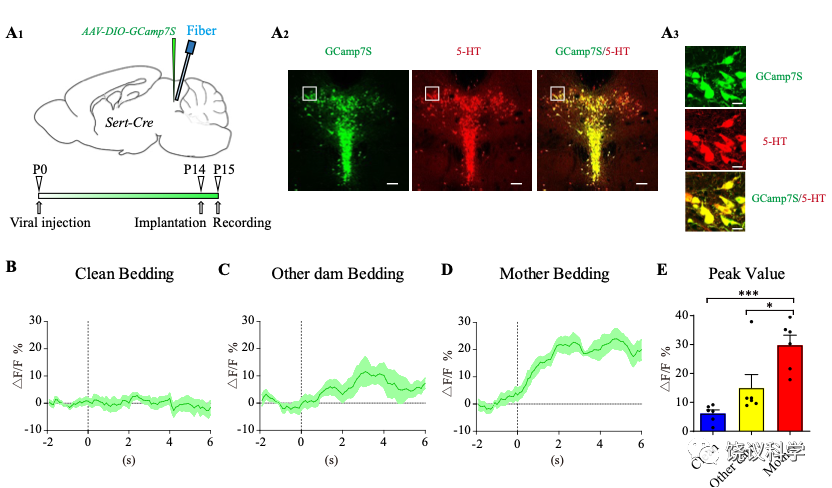
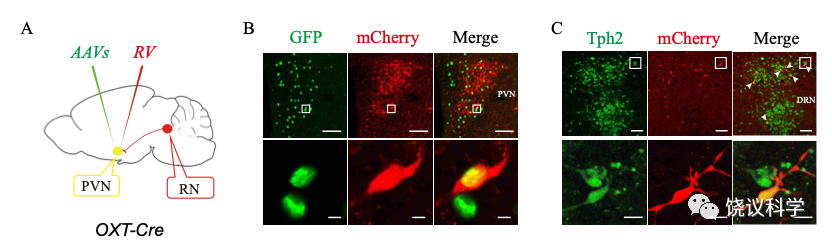
刘琰等把光导纤维插入小鼠脑内,并用病毒介导表达钙离子(Ca2+)敏感的蛋白质,通过特异控制元件指导其特异表达在中缝核(RN) 的五羟色胺能神经元(使用五羟色胺作为神经递质的神经元)。 他们因此可以通过钙离子浓度变化推测神经元的激活情况。他们发现,在给幼崽含母亲气味的垫料时,中缝核的五羟色胺能神经元被激活。而干净的垫料不激活、其他母亲气味的垫料引起的激活显著低于自己母亲气味垫料引起的激活。
用c-fos基因表达作为神经元激活的指示时,也可以观察到母亲气味激活中缝核五羟色胺能神经元。用c-fos方法还可以看到母亲气味激活室旁核的催产素能神经元。
缺乏催产素或催产素受体的大鼠幼崽有母亲偏好的缺陷、但无超声的缺陷
以前只有小鼠的催产素和催产素受体基因缺陷突变种,而其作用有争议。
刘琰等制造了大鼠的催产素和催产素基因剔除突变种。

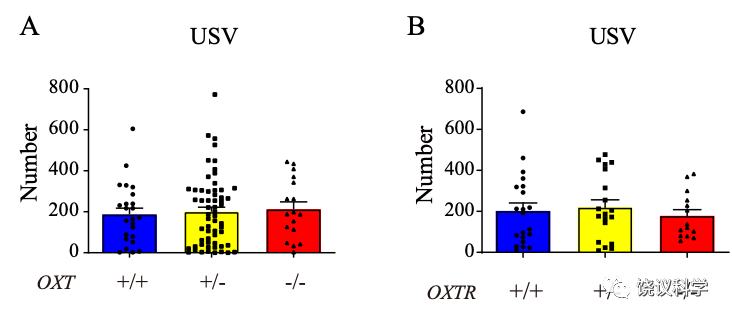
催产素基因剔除的OXT-/-大鼠幼崽与没有剔除的幼崽在母亲分离诱导的超声发放检测方面没有差别。催产素受体基因剔除也不影响这一检测结果。
在检测大鼠幼崽对母亲和干净垫料的偏好时,催产素和催产素受体的基因突变都没有影响。
但是,在检测大鼠幼崽对母亲和其他雌性垫料的偏好时,催产素和催产素受体都起显著作用。
这些结果表明,催产素和催产素受体参与亲母行为中的部分成分。
催产素和催产素受体不影响大鼠幼崽回避薄荷的行为。
亲母行为中五羟色胺在催产素上游起作用
分析突变动物显示,Tph2-/-, OXT-/- and OXTR-/- 大鼠幼崽有一个共同的变化: 影响对母亲和其他雌性气味的偏好。刘琰等进一步研究五羟色胺与催产素是否有关。
催产素是一种短肽,含有九个氨基酸。前人的研究表明它可以通过喷鼻而进入脑内。刘琰等给催产素基因剔除的大鼠幼崽喷鼻催产素,结果可以挽救催产素基因剔除大鼠幼崽的母亲气味偏好缺陷。而用另外一个九肽(加压素)没有这一作用。这两个九肽氨基酸序列很近,而作用不同显示了催产素在这一行为的特异性。
有趣的是,在缺乏五羟色胺的Tph2-/-基因剔除大鼠幼崽,催产素喷鼻也能使它们能够恢复对母亲气味的偏好。
以上实验结果提示催产素在五羟色胺的下游起作用。而给催产素基因剔除的大鼠幼崽注射五羟色胺合成前体5-HTP不能挽救其对母亲气味偏好的缺陷,进一步证明五羟色胺在催产素的上游调控幼崽对自己母亲的偏好行为。
催产素喷鼻不能挽救缺乏五羟色胺后大鼠幼崽在母亲诱导的超声发生缺陷,说明五羟色胺下游还有除外催产素的分子。
在用pCPA药理降低婴猴脑内的五羟色胺后,通过喷鼻给予催产素,可以挽救其与母亲待在一起的时间。说明从鼠到后,五羟色胺都在催产素上游发挥调控亲母行为的功能。
化学遗传学抑制背侧中缝核五羟色胺能神经元可以导致亲母行为缺陷
刘琰等用化学遗传学方法,将设计的受体基因 (DREADDS) 引入小鼠脑内特定神经元。然后给化学分子,通过不同的DREADDS,或抑制神经元、或激活神经元。通过病毒引入特定基因后,先验证所编码蛋白质的表达,再电生理记录验证确实能够抑制或兴奋特定神经元,最后检测行为。
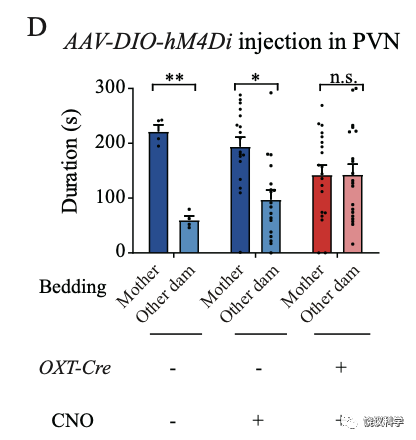
刘琰等抑制背侧中缝核的五羟色胺能神经元,发现小鼠幼崽因此出现亲母行为的缺陷。他们做了很多对照实验,排除是注射、病毒本身、药物本身等多种其他因素,确定不是其他原因而是因为抑制背侧中缝核的五羟色胺能神经元导致亲母行为缺陷。
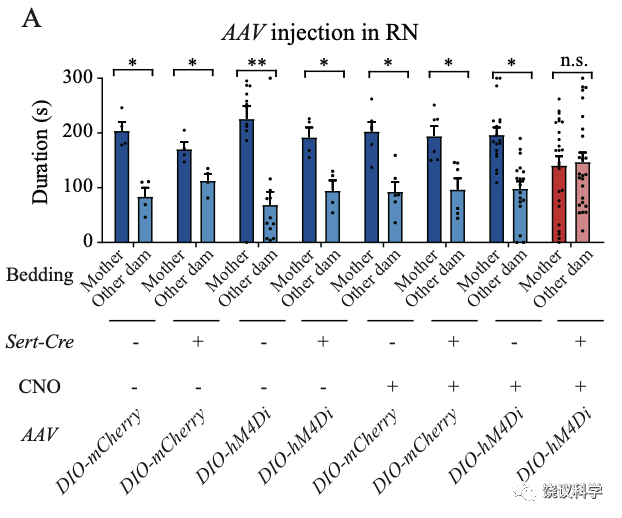
五羟色胺能神经元在背侧和腹侧中缝核都有,而且它们这两群都投射到室旁核。刘琰等通过实验区分了这两群,发现背侧中缝核参与亲母行为。
投射到室旁核的中缝核五羟色胺能神经元在亲母行为中的作用
已知中缝核投射到室旁核。刘琰等进行一系列实验,研究投射到室旁核的中缝核五羟色胺能神经元的作用。
刘琰等首先去除来自中缝核五羟色胺能神经细胞中的五羟色胺,在行为实验前,将表达Cre酶的病毒注射到Tph2-floxp基因敲入小鼠幼崽的中缝核。这样可以条件敲除Tph2基因:只有中缝核的Tph2基因被敲除,不产生五羟色胺。结果是亲母行为缺陷。
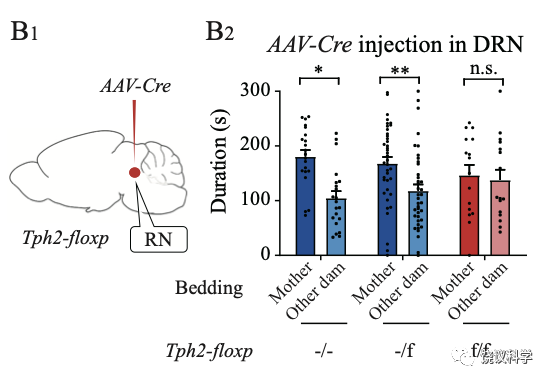
更为巧妙的是,刘琰等把表达Cre酶的病毒注射到Tph2-floxp基因敲入小鼠幼崽的室旁核(PVN),这样,只有投射到室旁核的、来自中缝核的五羟色胺能神经元的Tph2基因会被敲除。这时,小鼠幼崽也失去亲母行为。说明从中缝核投射到室旁核的五羟色胺能神经元中的五羟色胺对于亲母行为很重要。
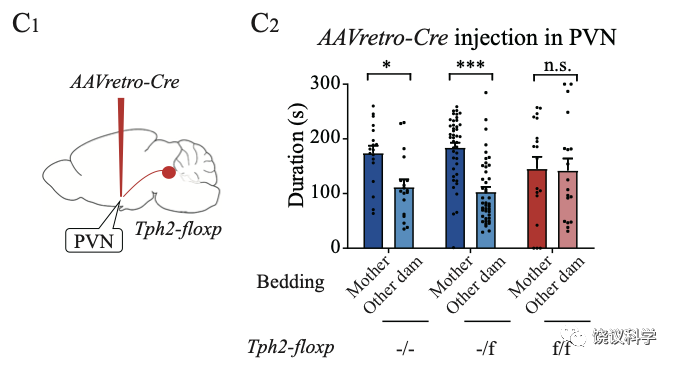
刘琰等用化学遗传学抑制室旁核的催产素能神经元,导致小鼠幼崽亲母行为缺陷。
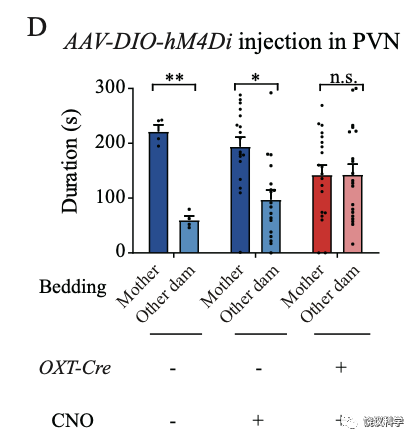
催产素能神经元在五羟色胺能神经元下游参与亲母行为的母亲偏好成分
刘琰等通过可以跨一个突触的病毒,证明室旁核的催产素能神经元直接接受中缝核来的五羟色胺能神经元的投射。
之后,他们通过实验证明这一投射的功能意义。
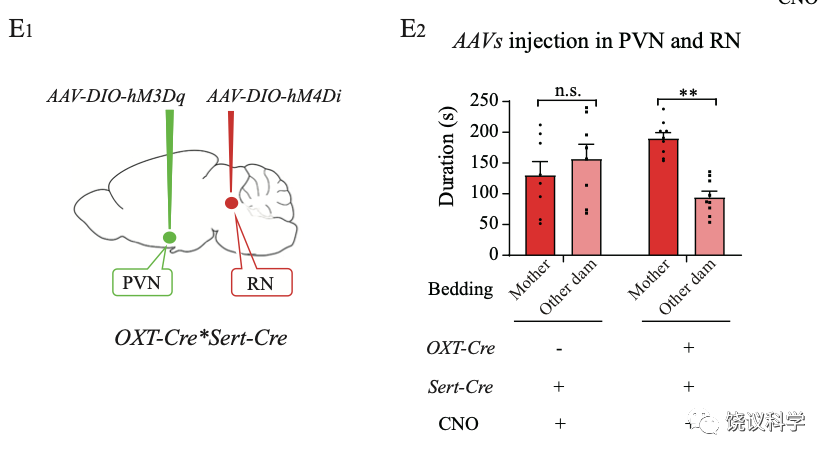
化学遗传学抑制室旁核的催产素能神经元活动后,小鼠幼崽缺乏母亲偏好。
抑制中缝核五羟色胺能神经元导致的母亲偏好缺陷,可以通过激活室旁核催产素神经元而得救。
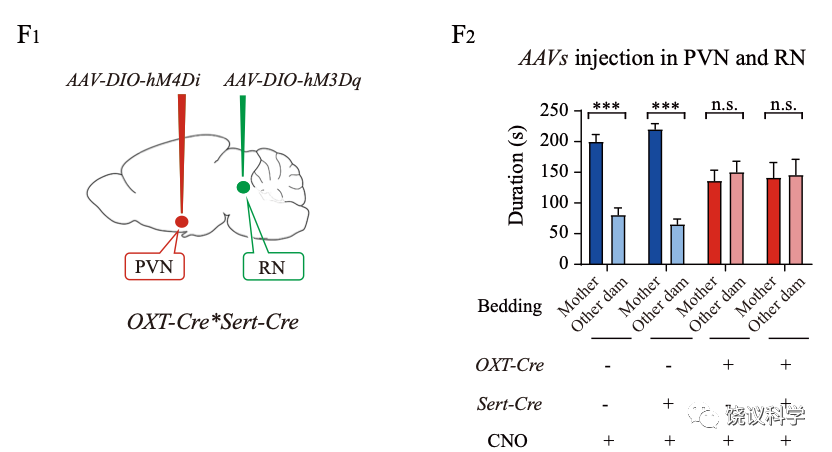
而抑制催产素能神经元导致的同样缺陷,不能通过激活中缝核五羟色胺能神经元得到挽救。说明催产素能神经元在五羟色胺能神经元下游发挥作用。
五羟色胺通过不同的受体,可以是激活、也可以是抑制下游神经元。刘琰等文章证明五羟色胺能够激活催产素能神经元。

光遗传学激活室旁核的五羟色胺能神经元末梢,也能够激活催产素能神经元。说明中缝核投射到室旁核的五羟色胺能神经末梢是激活室旁核催产素能神经元。
这些结果综合起来,说明中缝核的五羟色胺能神经元,投射神经末梢至室旁核,通过五羟色胺激活催产素能神经元,参与调控亲母行为的母亲偏好成分。
亲母行为是哺乳动物普遍存在的第一个社会行为。刘琰等通过多种动物的遗传和分子生物学操纵,证明五羟色胺对于亲母行为起着高度保守的重要功能。这些结果提示,但尚未证明,很可能五羟色胺对于人类婴儿的亲母行为也很重要。刘琰等的论文,是人类第一次同时报道小鼠、大鼠和猴的同一个基因的功能。
刘琰等的研究还综合了遗传学、分子生物学、药理学、电生理、光学成像、免疫组化、光遗传学、化学遗传学、神经生物学、行为学等多学科研究途径。
刘琰等的研究设计,经常是用较多的对照实验,严谨地对于一点、一点的结果进行反复验证。展示在8个主图、16个附图中。
这篇论文提出一个可能性,已经有很多实验证明催产素和加压素等神经肽在社会行为起重要作用,而饶毅实验室从2011年发表其第一篇有关哺乳动物中五羟色胺在性偏好的作用文章后,还发现五羟色胺对哺乳动物的多种社会行为很重要。在2023年这篇论文中,饶毅提出五羟色胺可能是哺乳动物的社会性神经肽上游的关键调控分子,分别通过多个下游分子,包括催产素和加压素,调控多个不同的社会行为、或同一社会行为的不同成分。
该论文发表在2023年3月2日上线的《神经元》杂志,题目是“第一个社会行为‘亲母’的分子和细胞机理:五羟色胺从鼠到猴起保守作用,其下游控制催产素”。
刘琰为第一和通讯作者,饶毅为通讯作者。其他作者为:单梁、刘田娥、李娟、孙常宏、卞希玲、席建忠、杨超娟、张晨、陈永昌、牛昱宇。
原文链接:https://www.cell.com/neuron/fulltext/S0896-6273(23)00114-9